بعد عرض أدلة التطور والرد على الطرح المضاد للتطور الذي يروج له الخلقيون، يحين موعدنا مع الرد على أشهر انتقادات وجهت لنظرية التطور.
التشابه ليس دليلًا على التطور
يعترض الخلقيون قائلين أن التشابه بين تراكيب الكائنات الحية ليس دليلًا على التطور. هذا ادعاء صحيح إلى حد ما، فليست كل التشابهات بين الكائنات الحية يمكن استخدامها على أنها دليل على تطورها من بعضها البعض بشكل مباشر.
على سبيل المثال، يمتلك كل من الطيور والحشرات الطائرة أجنحة، ولكن هذا لا يمكن أن يستخدم كدليل على انحدارهم من سلف مشترك امتلك أجنحة. في الواقع هذا ادعاء خاطئ، فقد طوّر كل منهم تركيبة الأجنحة بشكل منفصل. ولكن كيف يعرف العلماء ما إن كانت هذه التراكيب منحدرة من سلف مشترك أم تطورت بشكل منفصل؟
تكمن الإجابة في معرفة التركيبة والوظيفة للأعضاء التي نقارنها. فمثلًا، لدى كل من الخفافيش والطيور أجنحة، ولكنها مختلفة تمامًا في التركيبة على الرغم من تأديتهم لنفس الوظيفة. فعلى سبيل المثال، يتكون جناح الخفاش من غشاء يمتد بين أصابعه، بينما لا نرى ذلك في أجنحة الطيور.


ولكن عندما نأتي مثلًا لنرى زعانف الحيتان مقارنة بذراع الإنسان، نرى أنهما يتبعان نمطًا واحدًا كما ناقشنا في الجزء الأول من السلسلة. فعلى الرغم من امتلاكهم لوظائف مختلفة تمامًا، فنحن نرى نفس التركيبة. بالإضافة إلى أن زعانف الحوت لا تشبه زعانف الأسماك، ولكنها كما قلنا تتبع نمط الأطراف الأمامية لدى الثدييات. وهو ما يتوافق مع كونها ثدييات أصلًا.
باختصار، نوع التشابه الذي يستخدم للدلالة على التطور من سلف مشترك هو التشابه غير الضروري في تركيبة الأعضاء على الرغم من اختلاف الوظيفة. [1]
وجود فجوات في السجل الأحفوري
ينتقد الخلقيون نظرية التطور باعتبارها تفتقد لاكتمال السجل الأحفوري. فعندما يرون الكم الهائل من الأدلة التي تؤيد التطور من السجل الأحفوري، يقولون أن السجل الأحفوري غير مكتمل.
بالطبع هذا الادعاء صحيح، ولكنه لا يصلح لاستخدامه في نقد التطور. فتكوّن الحفريات عملية نادرة للغاية، ونحن على الأرجح لن نحصل على سجل أحفوري مكتمل في يوم من الأيام. ولكن هذا لا يصلح لمواجهة الكم الهائل من الحفريات التي عثرنا عليها، حيث أن كل حفرية من الحفريات الموجودة في السجل الأحفوري كانت “حلقة مفقودة” في مرحلة ما. على سبيل المثال، كان الاعتقاد سائدًا منذ زمن بعيد أن الإنسان والشيمبانزي متشابهان، ولكن لم تكن هناك حفريات انتقالية لتدعيم فكرة انحدار الإنسان والشيمبانزي من سلف مشترك. أصبح لدينا اليوم سجل أحفوري يزخر بالحفريات الانتقالية التي تحكي لنا قصة تطور الإنسان.
إذًا فمجرد وجود فجوات في السجل الأحفوري لن ينفي وجود عدد كبير من الحفريات الانتقالية المكتشفة بالفعل، ولا يصلح كنقد للتطور. يمكن اعتبار وجود فجوات في السجل الأحفوري دعوة مفتوحة للجميع للمشاركة في سد تلك الفجوات بممارسة البحث العلمي والحملات الاستكشافية لاكتشاف المزيد من الحفريات.
الانفجار الكمبري
ليس لدى الخلقيين حجة أفضل الانفجار الكمبري ليستخدموها في نقد التطور. إلا أنها مبنية على أساس نفس الادعاء السابق كما سنوضح. تتلخص حجتهم أن في مرحلة ما من تاريخ الأرض، ظهر عدد ضخم من الكائنات الحية بشكل “مفاجئ” في السجل الأحفوري. وأن داروين نفسه اعترف بندرة الحفريات مما قبل العصر الكمبري، مما يثبت فرضيتهم عن الخلق المباشر.
ولكن هذا ادعاء غير صحيح بالمرة، فالحياة لم تبدأ في العصر الكمبري. ولكن يمكننا اقتفاء أثر للحياة مما قبل الانفجار الكمبري بحوالي 3 بليون سنة! وصحيح أن الحفريات ما قبل الانفجار الكمبري كانت نادرة في عصر داروين، ولكن هذا كان في عصر داروين فقط!
وجد العلماء العديد من الحفريات مما قبل الكمبري، والتي تعتبر نماذجًا أولية لأشكال الحياة التي عاشت في العصر الكمبري. مثل «بارفانكورينا-Parvancorina»، والتي تُشبه «الترايلوبايت-Trilobites» التي عاشت في العصر الكمبري إلى حد كبير. ولا يمكننا معرفة ما إذا كانت هي الأسلاف الحقيقية للترايلوبايت، ولكننا نرى التشابه في الصفات التشريحية كما تتنبأ نظرية التطور بالضبط. كما عُثر عليها في نفس الطبقات الرسوبية التي يتنبأ التطور بوجودها فيها! [2]


كما عثر العلماء على «الكمبريلا-Kimbrella»، والتي تشابه الرخويات بدرجة كبيرة. ومجددًا، في نفس الطبقات الرسوبية التي تتنبأ بها نظرية التطور. [3] وغيرها الكثير من أشكال الحياة البدائية التي يمكننا العثور عليها مما قبل الانفجار الكمبري، وهذا يقودنا إلى استنتاج مفاده أن الانفجار الكمبري ليس “انفجارًا” حقًا.
أضف إلى علمك أن الانفجار الكمبري استمر لحوالي 20 مليون سنة، وهو ليس بالرقم الصغير! فمهما حدث من تنوع حيوي، فقد حدث على مدار 20 مليون سنة! [4]
في الواقع، لدينا أمثلة على تحولات تطورية كبيرة في فترات أقل بكثير من هذه، فلدينا سجل أحفوري جيد جدًا يوثق تطور الحيتانيات مثلًا. حتى أن نوعًا جديدًا من الحيتان البدائية اكتشف في وادي الحيتان في مصر في 2021 [5]، وأطلق عليه مكتشفوه اسم «فيوميسيتس أنوبيس-Phiomicetus Anubis». ونحن نعلم أن هذا التحول الكبير من حيوانات برية إلى الحيتان حدث في أقل من 10 مليون سنة فقط! [6]
إذًا فالعثور على حفريات انتقالية قبل الانفجار الكمبري تعد صورًا أولية للكائنات التي ظهرت فيه. ومعرفة أن الانفجار الكمبري استمر 20 مليون سنة، تكفي للإطاحة بحجة الانفجار الكمبري، وإثبات أنها لم تعد تصلح لاستخدامها في نقد التطور.
التطور يحدث داخل نفس النوع
يعترض الكثيرون قائلين أن التطور يحدث في نفس النوع فقط، ويتخيلون أن كل أمثلة التطور المرصود هي مجرد تكيفات ولا تصلح لاستخدامها كدليل على التطور. وللرد على هذا الادعاء علينا أولًا معرفة تعريف «النوع-species».
وفقًا للموسوعة البريطانية، النوع هو مجموعة من الكائنات الحية تضم أفرادًا يمتلكون خصائص متشابهة ويمكنهم التزاوج. [7]
والواقع أن هناك تجربة شهيرة توضح حدوث «الانتواع-Speciation»، مباشرة أمام أعيننا. حيث قامت «دايان دود-Diane Dodd» بتجربة بسيطة قامت فيها بعزل مجموعتين من ذباب الفاكهة، ووضعت كل مجموعة في قفص مغلق. قامت دايان بتغذية مجموعة منهم على طعام مكون بشكل أساسي من سكر «المالتوز-Maltose»، بينما غذّت المجموعة الأخرى بطعام مكوّن بشكل أساسي من «النشاء-Starch». وبعد فترة، أطلقت المجموعتين على بعضهما البعض لترى أنماط التزاوج التي سيتخذونها، وهنا كانت المفاجأة!
تزاوج ذباب المالتوز مع ذباب المالتوز، وذباب النشاء مع ذباب النشاء. هذا مثال على الانتواع يحدث أمام أعيننا، فقد أدى الانعزال والعيش في ظروف مختلفة لفترة من الزمن إلى ظهور نوعين من ذباب الفاكهة. [8] ولكنهم يقولون أن الذباب ظل ذبابًا، ولم يصبح أي كائن آخر. أجل، فهل تقول نظرية التطور أن الذباب سيبيض عصافيرًا مثلًا؟
ما يخبرنا به التطور أن التغيرات الصغيرة تتراكم على مدى زمني طويل لتصير تغيرات كبيرة ملحوظة. وكل التغيرات الصغيرة التي نرصدها في المعمل أو خارج المعمل ما هي إلا خطوات صغيرة إذا استمرت وتراكمت، ستصير الكائنات بشكل مختلف تمامًا. فتراكم السنتيمترات (على الرغم من صغرها) سيعطينا عاجلًا أم آجلًا كيلومترات، ولا يمكنك إنكار هذا لأنه لا يعجبك. وهنا قد تتساءل إذا لم يكن التطور الكبير مرصودًا فكيف للعلماء أن يعرفوا حدوثه؟
يعرف العلماء حدوثه من السجل الأحفوري الذي يوثق تطور أنواع كثيرة. كما يعرفونه من علم الأجنة الذي يوفّر أدلة قوية على التطور. وكذلك يعرفونه من الجينوم الذي أثبت حدوث التطور بأدلة قاطعة كما بينت كثيرًا في هذه السلسلة بطولها.
لوسي قرد عادي
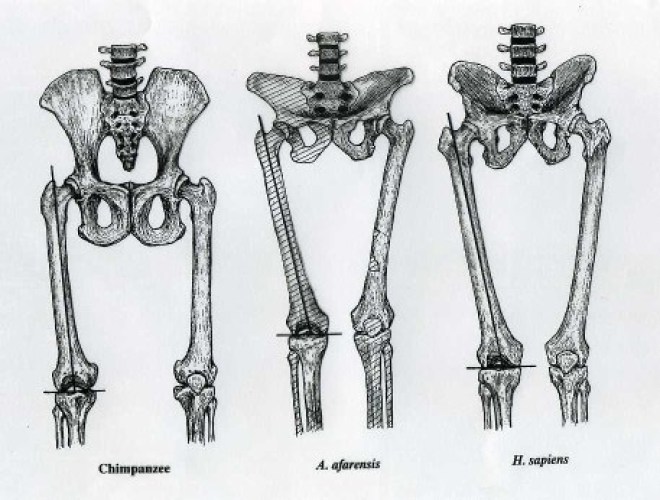
نعم، هذا صحيح تمامًا، فالتصنيف العلمي لي ولك وللشيمبانزي والغوريلا والأورانجوتان هو «القردة العليا-Great Apes». وكذلك كان الأوسترالوبيثيكاس (لوسي)، ولكن ليس هذا ما يعنيه الخلقيون، ما يقصدونه هو أن لوسي مجرد شيمبانزي وليست ضمن الخط التطوري للبشر.
وهذا غير صحيح لأننا نجد صفات بشرية لدى لوسي ليست لدى القرود غير البشرية، مثل المشي المنتصب. نعرف أن لوسي قد مشيت منتصبة عن طريق معرفة زاوية تمفصل عظمة الفخذ مع عظمة الحوض في الحفرية. كما عثرنا على حفريات لآثار أقدامها، وهي مشابهة لآثار البشر كما ناقشنا بتفصيل أكثر في الجزء الخاص بالسجل الأحفوري.
رسومات إرنست هيكل
يقولون أن خط الأدلة من علم الأجنة مبني على رسومات إرنست هيكل التي اكتشف لاحقًا أنها مزيفة. حيث رأى هيكل أن الجنين يمر بمراحل أثناء تكونه يكون مشابهًا فيها لأسلافه في مرحلة البلوغ، فيما يُعرف ب«التلخيص-Recapitulation» وهذا لأن الجنين -وفقًا لرأي هيكل- يُلخّص تاريخه التطوري أثناء مراحل تكونه. فعلى سبيل المثال، سيبدأ الجنين البشري بكائن أحادي الخلية، ثم سمكة، ثم يصير كائنًا برمائيًا، وهكذا إلى أن يصل إلى الهيئة البشرية (وفقًا لفرضية هيكل).
وهذا مجرد هراء، فقد قمنا بتخطئة نظرية هيكل في أول فقرة من مقال علم الأجنة. كما دعّمنا الأدلة التي أوردناها من علم الأجنة بصور فوتوغرافية لا علاقة لها برسومات هيكل من قريب أو بعيد. وقد تجاوز العلم رسومات هيكل بسنوات من البحث والتدقيق والتمحيص. ولكن يبقى الخلقيون مولعون بالبحث في ركام الفرضيات العلمية التي قام المجتمع العلمي نفسه برفضها لينتقدوا بها العلم الحديث، وفي هذا تدليس واضح.
عدد كروموسومات الإنسان
يردد الخلقيون عن جهل هذا المثال كثيرًا، فيقولون كيف للإنسان أن يمتلك عددًا مختلفًا من الكروموسومات عن القردة العليا إن كان منهم؟ والحقيقة أن الكروموسوم 2 لدى الإنسان عبارة عن كروموسومين ملتحمين كما أثبتنا بشكل أكثر تفصيلًا في مقال أدلة الجينوم. هذا بغض النظر عن وجود حالات مرضية لتغير عدد الكروموسومات داخل نفس النوع، فلدينا نحن البشر مجموعة من التغيرات الكروموسومية بالفعل:
• «متلازمة داون-Down syndrome»، وتنتج عن وجود كروموسوم زائد مماثل للزوج رقم 21، وبالتالي فإن كل خلايا الجسم تحتوي على 47 كروموسوم بدلًا من 46.
• «متلازمة كلاينفلتر- Klinefelter syndrome»، وتنتج عن وجود كروموسوم X زائد في الحيوان المنوي، فتصبح كروموسومات الجنين XXY.
• «متلازمة تيرنر-Turner syndrome»، وتحدث في الإناث نتيجة عدم نقص وجود كروموسوم X بشكل كلي أو جزئي. [9]
كما أن الأبحاث العلمية أثبتت بشكل قطعي أن الكروموسوم 2 عبارة عن كروموسومين ملتحمين، لذا فلا فائدة من نكرانه أو محاولة الهروب منه. فليتعامل الخلقيون مع ذلك!
«سمكة السيليكانث-Ceolacanth»
يحتج الخلقيون بأن سمكة السيليكانث لم تتطور على مدى ملايين السنين وهذا يجعل منها «أحفورة حية-Living fossil»، وأن هذا يعارض التطور. وهو ادعاء باطل من وجهين:
أولًا، لا تحتم نظرية التطور على كل الكائنات أن تستمر في التغير، فالانتخاب الطبيعي يحافظ على الصفات المناسبة لبيئتها بشكل كافٍ. لذا فوجود أي كائن لم يتغير على مدى سنين كثيرة لا يُعد حجة لتستخدم في نقد التطور.
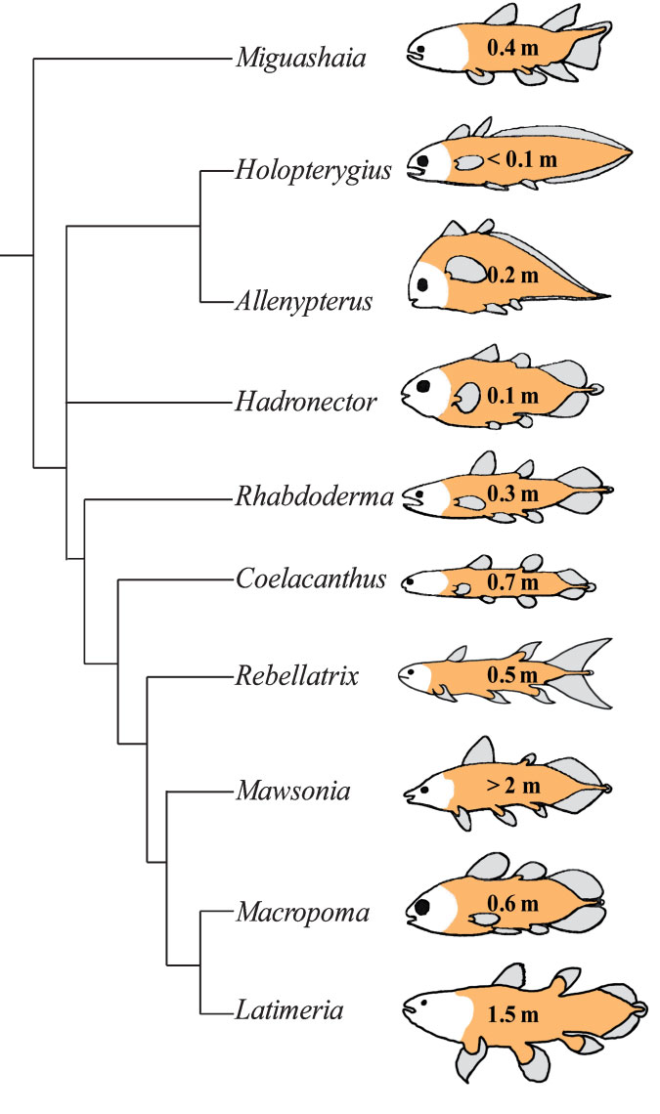
ثانيًا، لا تعد السيليكانث أحفورة حية أصلًا، فالسجل الأحفوري يزخر بأقرباء السيليكانث المنقرضين مثلما نرى في هذه الصورة (سمكة Latimeria هي نوع السيليكانث الموجود حاليًا). [10]

منذ أن خرجت نظرية التطور إلى النور وهي مبحث مهم من مباحث العلم الحديث. لذا فقد رأينا أن نعرض لكم أبرز أدلة التطور وأكثرها شيوعًا، مع العلم بأن هناك عدد أكبر من الأمثلة في كل خط من خطوط الأدلة التي ناقشناها. ويستمر البحث في نظرية التطور، وتستمر الأدلة في الزيادة، ونستمر في إطلاعكم بكل جديد في مجال العلوم.
المصادر
[1] Berkeley
[2] Fossil Fandom
[3] Nature
[4] Springer
[5] Royal Society
[6] Whales
[7] Britannica
[8] Berkeley
[9] About Kids Health
[10] Ecologica




























































/2048px-Archaeopteryx_fossil-5c5b17f246e0fb0001849b0e.jpg?w=660&ssl=1)