- الأدلة السلوكية على التطور
- الأدلة على التطور من علم الأجنة
- أدلة التطور: مشاهدة التطور في المعامل
- لماذا تسمى نظرية التطور وليس قانون التطور؟
- الأدلة التشريحية على التطور
- أدلة التطور: الأعضاء الضامرة
- أدلة التطور من السجل الأحفوري
- أدلة التطور: التطور السريع
- أدلة تطور الحيتانيات
- أدلة التطور من الجينوم
- التصميم الذكي في ميزان العلم
- ردود على نقد التطور
- اختبار مقدمة في نظرية التطور
في مقال اليوم نتعرف على أقوى خط من خطوط أدلة التطور، إذ سنتحدث عن أدلة التطور من الجينوم. الأدلة الجينومية هي نوع حديث نسبيًا من أنواع الأدلة على التطور، حيث أن كل ما سنذكره في مقال اليوم من أدلة لم يكن معروفًا قبل القرن الماضي. لقد تطورت علوم الجينوم بقوة وأصبحت ركيزة أساسية في علوم الطب والصحة والبيولوجيا. توقع الكثيرون أنها ستوجه ضربة قاضية لنظرية التطور، ولكنها على العكس، وجهت ضربة قاضية لكل ما يخالف التطور. لقد ظهرت جينات الأعضاء الضامرة بوضوح، بل وكيف ضمرت جينيًا. أصبح بالإمكان رسم شجرة التطور بواسطة الحواسيب من خلال تحليل تتابعات متعدد لجين معين ومعرفة رحلته التطورية عبر الكائنات الحية. والآن لنستعرض الأدلة، ولكن علينا أولًا أن نفهم بعض أساسيات علم «الأحياء الجزيئية-Molecular Biology».
محتويات المقال :
ما هو ال DNA؟
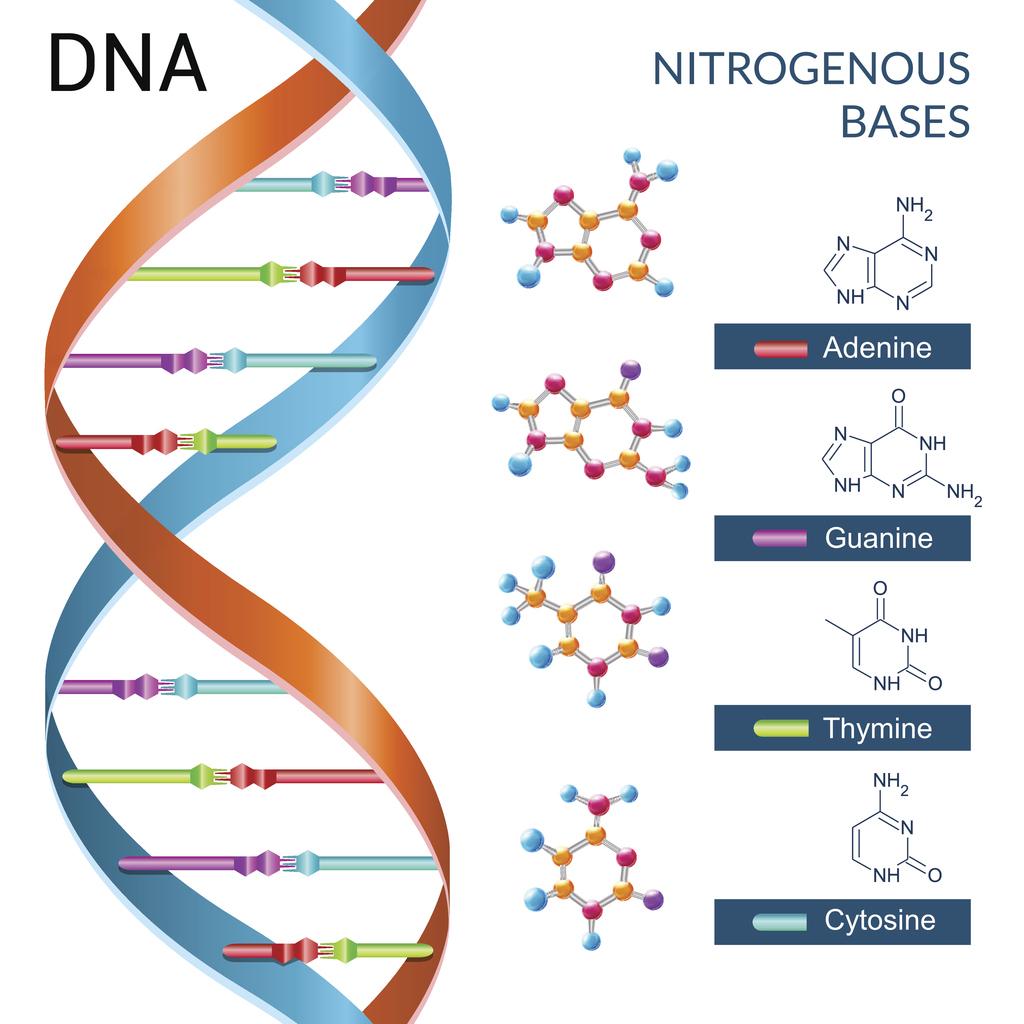
تتكون كل الكائنات الحية التي تعيش على كوكبنا من خلايا، فالخلية هي الوحدة الأساسية لبناء الكائن الحي. وفي نواة الخلية، نجد جزيئات معقدة تُدعى «الحمض النووي الريبي منقوص الأوكسجين-Deoxyribonucleic acid» وهي ما نرمز له ب “DNA”.
ويتكون ال DNA من تسلسل حروف هي A,C,T,G وكل حرف منهم يرمز إلى مركب كيميائي، حيث أن:
• حرف A يرمز إلى «أدينين-Adenine»
• حرف C يرمز إلى «سيتوزين-Cytosine»
• حرف T يرمز إلى «ثيامين-Thymine»
• حرف G يرمز إلى «جوانين-Guanine»
وتُدعى هذه المركبات الأربعة ب «القواعد النيتروجينية-Nitrogenous bases». عند اتحاد القواعد النتيروجينية مع جزيء «سكر خماسي الكربون-Pentose sugar»، و«مجموعة فوسفات-Phosphate group» تكون «النيوكليوتيدات-Nucleotides»، وهي وحدة البناء الأساسية لل DNA.
ما هي الجينات؟ وكيف تعمل؟
الجين هو قطعة لها بداية ونهاية من شريط الDNA، بمعنى أدق، هو تسلسل محدد من الحروف A,C,T,G. يحمل هذا التسلسل الأوامر اللازمة لإنتاج بروتين محدد، كأنها حروف في كتيب تعليمات لتركيب أجزاء المروحة. وفقًا لترتيب تلك الحروف، تترتب الأجزاء معًا وينتج في النهاية مروحة تعمل بكفاءة. ولكن كيف تعمل الجينات؟
يتم نسخ تسلسل حروف الجين الموجودة على شريط الDNA إلى تسلسل حروف آخر يُسمى «RNA رسول-Messenger RNA» ويُرمز له ب”mRNA”. ينتقل جزيء الmRNA هذا من نواة الخلية إلى «الريبوسوم-Ribosome» (أحد العُضيّات الموجودة داخل الخلية)، ويعمل الريبوسوم كآلة لإنتاج البروتين، حيث يقوم بقراءة تسلسل حروف الmRNA. تعطي كل 3 حروف الأمر لاختيار أي نوع من أنواع «الأحماض الأمينية-Amino Acids» بالتحديد ليضعه الريبوسوم في السلسلة. على سبيل المثال، الحروف “CAA” تخبر الريبوسوم أن يختار «الجلوتامين-Glutamine» (أحد الأحماض الأمينية). وعند الانتهاء من قراءة الmRNA تكون عملية تكوين البروتين قد تمت، ويمكنك مشاهدة مقطع يوضح هذه العملية الرائعة من هنا. [1]
وبعد هذه المقدمة التمهيدية البسيطة، ننتقل إلى استعراض أدلة التطور من الجينوم.
وحدة الشيفرة الجينية
كما ذكرنا، تستخدم كل الكائنات الحية على الأرض الDNA لتشفير معلوماتها الوراثية. كما تستخدم نفس طريقة صنع البروتين، حيث تعطي نفس الثلاث حروف الأمر للريبوسوم لوضع نفس الحمض الأميني في سلسلة الأحماض الأمينية لصنع البروتين في كل الكائنات الحية، ولكن ما هو تفسير هذا؟
وقد يتساءل القارئ عما إن كان هذا يتطلب تفسيرًا أصلًا، ولكن صدق أو لا تصدق عزيزي القارئ، شيفرة الDNA ليست الشيفرة الوحيدة الممكنة.
أظهرت الأبحاث باستخدام برامج الحاسوب إمكانية تشفير المعلومات الوراثية بما يزيد عن مليون شيفرة جينية ممكنة غير الDNA. بل أن بعضها قد يؤدي وظيفته بشكل أفضل بكثير من الDNA، فلماذا تستخدم كل الكائنات نفس الشيفرة؟ [2] , [3]
لا يمكننا فهم هذا إلا في ضوء نظرية التطور من أصل مشترك. فنظرية التطور تخبرنا أن جميع هذه الكائنات ورثت شيفرتها الوراثية من نفس الكائن الأولي.
ولا يمكننا أن نفهم سر وحدة الشيفرة في ضوء فكرة الخلق الخاص. لو كانت كل الكائنات قد خُلقت بشكل منفصل لما كان هناك حاجة لتكون هناك شيفرة موحدة. بل على العكس، فوجود شيفرات مختلفة في الكائنات المختلفة كان ليمنع انتقال الفيروسات بين الحيوانات المختلفة، مثل إنفلونزا الخنازير، وإنفلونزا الطيور والكورونا التي تصيب البشر، وغيرها من الفيروسات.
نسبة التشابه الجيني بين الرئيسيات
لا تتشارك الكائنات نفس الشيفرة فحسب، بل تتشارك في كمية كبيرة من الجينات. وتختلف نسبة التشابه الجيني بين الكائنات وفقًا لقربها أو بعدها عن بعضها البعض تطوريًا.
تبلغ نسبة الاختلاف الجيني بين البشر نسبة ضئيلة تُقدر ب 0.1%، وتستخدم نسبة التشابه في معرفة الأنساب وتحديدها قضائيًا اليوم. أما بيننا وبين أولاد عمومتنا من الشيمبانزي وقرود البونوبو تبلغ نسبة التشابه حوالي 1.2%، بينما تصل إلى 1.6% بيننا وبين الغوريلا. والأهم من ذلك، هو أن كل من البشر والشيمبانزي والبونوبو يتشاركون نفس نسبة الاختلاف الجيني عن الغوريلا. أي أن الشيمبانزي والبونوبو أقرب إلى الإنسان منهم إلى الغوريلا.
وتختلف طرق حساب نسبة التشابه والاختلاف الجيني، وباختلاف الطريقة تختلف النسبة. حيث أنه بأخذ الجينوم الكامل بالحسبان، تُظهر النتائج أن هناك مناطق من الDNA قد حُذفت، أو تمت مضاعفتها مرات ومرات. أضيف بعضها في أماكن أخرى من الجينوم أيضًا. وعند الأخذ بكل هذه الاختلافات، قد تصل نسبة الاختلاف الجيني بين الإنسان والشيمبانزي إلى ما بين 4% و5%. ومهما اختلفت طرق الحساب فالنتيجة واحدة، وهي أن الإنسان والشيمبانزي والبونوبو أقرب إلى بعضهم منهم إلى الغوريلا أو أي من الرئيسيات الأخرى. وهذا يخبرنا بحقيقة مدهشة وبديهية في الأوساط العلمية اليوم، وهي أننا لسنا فقط أقارب القردة العليا، بل نحن منهم!
وتتيح لنا دراسة نسب التشابه الجيني بين الكائنات بناء شجرة عائلة تربط هذه الكائنات ببعضها البعض. والمفاجأة أن هذه الشجرة تطابق ما يخبرنا به كل من التشريح المقارن ودراسة الحفريات في ضوء نظرية التطور. [4]

يعترض بعض المشككين قائلين أن التشابه الجيني لا يعني وحدة الأصل. لكن الواقع يقول أن الجينات لا تنتقل بين الكائنات الحية إلا بالتكاثر، فكون الكائنات تمتلك نفس الجينات يعني انحدارها من نفس الأصول. وكما تثق في تحليل الDNA الذي يخبرك إذا ما كان الطفل ابنك البيولوجي أم لا، عليك أن تثق بنتائج تلك الأبحاث التي تخبرك عن علاقتك التطورية بينك وبين أبناء عمومتك من الشيمبانزي والبونوبو وباقي الرئيسيات. علمًا بأن تحليل الDNA يتم بنفس الطرق التي قيست بها هذه النسب.
الكروموسوم 2
لدى الإنسان 23 زوجًا من الكروموسومات (46 كروموسوم)، بينما تمتلك القردة العليا الأخرى (الشيمبانزي، والأورانجوتان، والبونوبو، والغوريلا) 24 زوجًا (48 كروموسوم). غالبًا ما يلجأ المشككون لهذه المعلومة لدحض التطور، فيقولون إذا كان الإنسان من القردة العليا، فلماذا يختلف عنهم في عدد الكروموسومات؟
وتكمن الإجابة في أن تسلسل الDNA على الكروموسوم 2 لدى الإنسان متطابق تقريبًا مع كروموسومين منفصلين لدى الشيمبانزي. اقترح العلماء أن سبب اختلاف عدد الكروموسومات وفقًا لنظرية التطور هو أن الكروموسوم 2 عبارة عن كروموسومين مندمجين من كروموسومات القردة العليا، ولكن كيف نختبر هذه الفرضية؟
تُدعى أطراف الكروموسومات «تيلوميرات-Telomeres»، كما أن هناك منطقة تقع في منتصف الكروموسومات تُدعى «سنترومير-Centromere». يحتوي الكروموسوم الطبيعي على 2 تيلومير (نهايتين) وسنترومير واحد. فإذا كان الكروموسوم 2 في الإنسان عبارة عن اندماج في كروموسومين في القردة العليا، فعلينا أن نعثر على آثار لهذا الالتحام.
وجد العلماء آثار هذا الالتحام بالفعل، فقد أظهرت الأبحاث وجود بقايا تيلومير في منتصف الكروموسوم 2 (ناتج عن التحام تيلوميري الكروموسومين)، وبقايا سنترومير كذلك. هذا ما يثبت أن الكروموسوم 2 هو بالفعل عبارة عن اندماج كروموسومين من كروموسومات القردة العليا بشكل قاطع ولا يمكن تفسيره إلا باندماج كروموسومين. يدل هذا بما لا يدع مجال للشك على أصولنا المشتركة مع القردة العليا الأخرى. [5] , [6]

تكرار CMT1A الأقرب
على الكروموسوم 17 في الإنسان يقع جين PMP-22 الذي يشفر لإنتاج بروتين (Peripheral nerve protein-22)، محاطًا بنسختين من جين CMT1A، وهما «تكرار CMT1A الأقرب-Proximal CMT1A-REP»، و«تكرار CMT1A الأبعد-Destal CMT1A-REP».
ويؤدي «تعابُر الكروموسومات-Chromosome crossover» غير المتساوي، الناتج عن سوء اصطفاف التكرارين الأبعد والأقرب للجين CMT1A إلى مرضين عصبيين شائعين هما:
• Charcot-Marie-Tooth disease type (1A) (CMT1A)
• Hereditary Neuropathy with liability to Pressure Palsies (HNPP).
وبهذا نعتبر وجود نسختين من جين CMT1A خطأً وراثيًا، إذ يتسبب هذا التكرار غير الضروري بالإصابة بالأمراض العصبية، ولكن كيف يدل هذا على التطور؟
أظهرت تحاليل الDNA للرئيسيات امتلاكها جميعها لتكرار CMT1A الأبعد، ولكن فقط الإنسان والشيمبانزي وحدهما يمتلكان تكرار CMT1A الأقرب جنبًا إلى جنب مع التكرار الأبعد. أي أن هذا الخطأ الوراثي موجود فقط في الإنسان والشيمبانزي، ويمكننا تفسير هذا بحدوث عملية تضاعف للجين CMT1A في آخر سلف مشترك للإنسان والشيمبانزي.
لا يمكننا تفسير وجود هذا الخطأ لكل من الشيمبانزي والإنسان إلا وفق نظرية التطور. إذ ستتضاءل احتمالية حدوث نفس الخطأ لدى كل من النوعين بشكل منفصل دونًا عن بقية الرئيسيات التي أجريت عليها التحاليل. يصبح الأمر منطقيًا وفقًا لشجرة العائلة التطورية للقردة العليا. [7]
«الجينات الزائفة-Pseudogenes»
أثناء عملية التطور، تحدث طفرات لبعض الجينات متسببة في تعطيلها وتحويلها إلى جينات غير مشفرة للبروتين. تسمى هذه الجينات بالجينات الزائفة، لأنها لا تؤدي وظيفتها. تعتبر تلك الجينات الزائفة من أوضح الأمثلة على نظرية التطور، إذ لا يوجد ما يفسرها سوى النظرية. فهل من أمثلة؟
تنتج أجسام جميع الثدييات فيتامين C. إلا أن أجسام «الرئيسيات جافة الأنف-Haplorhini» (مجموعة تشمل قرود العالم القديم، وقرود العالم الجديد، والقرود العليا ومنها الإنسان) لا تستطيع صنع فيتامين C. تحصل الرئيسيات جافة الأنف علي فيتامين سي من غذائها، وفي حال عدم حصولها على الكميات الكافية، تُصاب بداء «الأسقربوط-Scurvy». يرجع هذا إلى عدم قدرتها على تصنيع إنزيم “l-gulono-gamma-lactone oxidase”، نظرًا لتعطل جين GULO المسئول عن إنتاج هذا الإنزيم.
المثير في الأمر هو أن جين GULO معطّل في جميع الرئيسيات جافة الأنف لنفس السبب، وهو فقدان «الإكسونات-Exons» (نطاقات مشفرة في الجينات) أرقام 1,2,3,5,6,8,11. وهو ما لا يمكننا فهمه إلا في ضوء انحدار الرئيسيات جافة الأنف من سلف مشترك.
ولا تصمد فكرة الخلق الخاص أو التصميم الذكي في محاولة تفسير الجينات الزائفة. فلا داعي لوجود جين GULO في تلك الأنواع إن كان معطلًا أصلًا. كما أنه لا داعي لأن يتم تعطيل الجين بفقدان نفس الإكسونات بين مختلف أنواع الرئيسيات جافة الأنف. إذ أن نفس الجين معطل في «خنزير غينيا-Guinea pig»، ولكن لأسباب مختلفة وهي فقدان الإكسون 5 وجزء من الإكسون 6. مما يعني أن هذا الجين قد تعطل لديه بشكل منفصل عن المسار التوري للرئيسيات جافة الأنف التي ينتمي إليها البشر. وبالتالي، لا يمكن تفسير تعطل جين GULO إلا في ضوء نظرية التطور من سلف مشترك. [8]
وكذلك حيوان «البلاتيبوس-Platypus»، الذي لا يمتلك معدة حقيقية، وعلى الرغم من ذلك، يمتلك جينين زائفين مرتبطين بصنع إنزيمات هضمية. [9]
وعلى الرغم من عدم امتلاك الحيتان البالينية للأسنان، فهي تمتلك جين MMP20 معطل يشفر لصنع بعض البروتينات التي تدخل في تركيبة الأسنان. وكما ذكرنا في الجزء الخاص بتطور الحيتانيات، تنمو لدى أجنة الحيتان البالينية براعم أسنان بالفعل. [10] كما تمتلك الطيور (وهي حيوانات بلا أسنان) جينات معطلة لتكوين أسنان! [11]
ووجود هذه الجينات الزائفة يعد من أقوى أدلة التطور من الجينوم، كما أنه دعم الأدلة التشريحية للتطور.
كيس المُح
نشترك نحن والزواحف والدجاج في امتلاكنا للبيض (البويضة في حالة إناث البشر). إلا أن الطيور (مثل الدجاج) لديها قشرة تحمي البيضة، كما تمتلك كيس المُح، والذي يقوم بتوفير الغذاء للجنين لمساعدته على النمو. ولكن بويضة الإنسان أصغر بكثير من بيض الدجاج، وليس فيها قشرة أو مُح حتى. ولكن تكون الأجنة البشرية كيس مح فارغ يختفي أثناء فترة الحمل، فهل من تفسير جيني؟
تمتلك كل الفقاريات التي تبيض بروتين Vitellogenin، مما يعني وجود جين يشفر لصنع هذا البروتين، فماذا تظهر الأبحاث؟
تظهر الأبحاث وجود قطع من DNA المُح في الجينوم البشري، مطابقة تماما للجينات الأصلية التي لدى الدجاج. ليس هذا وحسب، بل إنه عند مقارنة جينوم الإنسان بجينوم الدجاج يتضح أن هذه القطع محاطة بنفس الجينات لدى كل من الإنسان والدجاج. يعد هذا أحد أدلة التطور من الجينوم. [12] , [13]
جين NANOG
جين NANOG هو أحد «الجينات الزائفة المعالَجة-Processed pseudogenes». وهي جينات تنشأ نتيجة خطأ أثناء عملية نسخ حروف الDNA إلى mRNA. فبدلًا من أن تكتمل عملية إنتاج البروتين بشكل طبيعي، يُعاد بالخطأ نسخ الmRNA إلى الDNA لتكوين نسخة زائفة من الجين الأصلي في مكان آخر في الجينوم. هذا ما حدث مع NANOG، حيث نجد على الجينوم الخاص بنا 10 نسخ زائفة من هذا الجين بالإضافة إلى النسخة الأصلية التي تعمل. فكيف يكون هذا دليلًا على التطور؟
تكمن المفاجأة في أننا نتشارك 9 نسخ زائفة من جين NANOG مع أبناء عمومتنا من الشيمبانزي. بينما النسخة NANOGP8 حصرية لدى جينوم الإنسان. والأدهى، أن هذه النسخ ال 9 موجودة على جينوم الشيمبانزي في نفس الأماكن التي توجد فيها على جينوم الإنسان! إن احتمالية حدوث نفس الأخطاء 9 مرات في نفس المكان لدى النوعين بشكل منفصل احتمالية غاية في الضآلة. وأن تصدف هذه الاحتمالية شديدة الضآلة بين كائنين تصفهم نظرية التطور بالأقرابة يصعب الأمر على منكرين التطور. المشكلة لدى منكري التطور هو في إيجاد تفسير آخر خارج نظرية التطور لتلك الأدلة الحاسمة.
وهذا يعني أننا ورثنا 9 نسخ NANOG زائفة من آخر سلف مشترك بيننا وبين الشيمبانزي، أما عن النسخة المتبقية وهي NANOGP8، فهي وجدت في جينوم الإنسان بعد انفصاله عن الشيمبانزي، وهذا يعد أحد أقوى أدلة التطور من الجينوم. [14]
«سيتوكروم سي-Cytochrome C»
هناك جينات تتشاركها كل الكائنات الحية على اختلافها، لأنها تؤدي وظائف أساسية للغاية. جينات تتشاركها كل الكائنات ابتداء من أشكال الحياة وحيدة الخلية وحتى الإنسان، وتُعرف هذه الجينات ب «الجينات واسعة الانتشار-Ubiqitous genes».
وهذه الجينات ليس لها دخل بالصفات الخاصة بكل نوع، إذ أنها كما قلنا أعلاه، مسئولة عن وظائف أساسية للغاية تتشاركها معظم أشكال الحياة. ولا يُشترط أن تكون هذه الجينات بنفس تسلسل الحروف، إذ أن هناك عدد كبير من التسلسلات التي تؤدي نفس الوظيفة الحيوية الكيميائية.
والوراثة كما ذكرنا في أمثلة كثيرة أعلاه، تسمح بتمرير تسلسلات الجينات حتى وإن لم يكن لهذه التسلسلات فائدة وظيفية. وعلى هذا، فلا يمكننا تفسير وجود نفس التسلسل في هذه الجينات بين نوعين إلا بالوراثة؛ ذلك لأنهم لا يحتاجون نفس التسلسل إن كان كل منهم قد وجد بشكل منفصل كما يدعي المنكرون. يكفي وجود جين مكافئ يؤدي وظيفة مشابهة وهو أمر يحدث في الطبيعة بالفعل.
على سبيل المثال، نحن نعلم أن الإنسان والشيمبانزي متشابهين في كثير من الصفات. فيمكننا البناء على هذا، وتنبؤ استخدامهم لبروتينات متشابهة، بغض النظر عن ما إذا كانت تجمعهم علاقة وراثية أم لا. يصبح الأمر أكثر غرابة عندما نقارن بين تسلسلات جينية غير مؤثرة على خصائص أي من النوعين، فعلام نتحدث؟
نحن نتحدث عن بروتين «سيتوكروم سي-Cytochrome C»، وهو بروتين واسع الانتشار، إذ يوجد في كل أنواع الكائنات الحية بما في ذلك البكتيريا. وهو موجود في الميتوكندريا، حيث يقوم بنقل الإلكترونات في عملية أيض أساسية تُدعى ب «الفسفرة المؤكسدة-Oxidative phosphorylation».
ومما سبق يمكننا استنتاج أن امتلاك كل الكائنات لنفس تركيبة سيتوكروم سي شيء غير ضروري بالمرة، إذ يكفي فقط امتلاكها لبروتين مكافئ يؤدي نفس الوظيفة.
كما أظهرت الأبحاث أن سيتوكروم سي الخاص بالإنسان يعمل لدى الخميرة التي تم حذف نسخة سيتوكروم سي الخاصة بها. على الرغم من أن نسبة الاختلاف بين تركيبة السيتوكرومين تبلغ 40%.
في الواقع، سيتوكروم سي الخاص بالتونا (أسماك)، والحمام (طيور)، والأحصنة (ثدييات)، وذباب الفاكهة (حشرات)، والفئران (ثدييات)، جميعها تعمل في الخميرة التي تم حذف سيتوكروم سي الخاص بها!
علاوة على ذلك، أثبت تحليل جيني شامل لسيتوكروم سي أن أغلب الأحماض الأمينية الموجودة في تسلسل هذا البروتين غير ضرورية. وأنه فقط حوالي ثلث الأحماض الأمينية ال100 الموجودة في سيتوكروم سي كافية للقيام بوظيفته، وذلك لأن معظم الأحماض الأمينية في سيتوكروم سي هي أحماض «مفرطة التغيير-Hypervariable». والأحماض المفرطة التغيير يمكن استبدالها بعدد كبير من الأحماض الأمينية الأخرى التي ستكون قادرة على إتمام نفس الوظيفة.
تجربة هوبرت
والغريب في الأمر هو قيام الفيزيائي Hubert P. Yockey بحساب عدد تسلسلات الأحماض الأمينية الممكنة لتكوين البروتينات المكافئة لسيتوكروم سي، والقادرة على القيام بنفس تلك الوظيفة الأساسية. تبيّن أن عدد هذه التسلسلات الممكنة هو 10⁹³ × 2.316، وهو عدد مهول يزيد عن عدد الذرات في الكون المعروف بأكثر من بليون مرة! ومما سبق نستنتج أن تشابه تركيبة سيتوكروم سي بين الكائنات ليس ضروريًا على الإطلاق، فهل يخبرنا المنكرون سبب حدوثه إن كانت نظرية التطور خاطئة؟
ولكن العجيب هو أن الإنسان والشيمبانزي يتشاركون نفس تركيبة سيتوكروم سي، بلا أي اختلاف، واحتمالية حدوث هذا إذا أنكرنا العلاقة الوراثية بين الإنسان والشيمبانزي أقل من 10^-93 (1 من 10⁹³)، ويختلف كل من الإنسان والشيمبانزي عن باقي الثدييات الأخرى ب10 أحماض أمينية في تسلسل سيتوكروم سي فقط، وفرصة حدوث هذا إذا أنكرنا علاقتهم التطورية هي 10^-29 (1 من 10²⁹).
والخميرة أحد أبعد الكائنات «حقيقيات النوى-Eukaryotic» عن الإنسان، إلا أنها تختلف عنّا في تركيبة سيتوكروم سي ب51 حمض أميني فقط، وفرصة حدوث هذا دون وجود علاقة وراثية بيننا وبين الخميرة أقل من 10^-25 (1 من 10²⁵). [15]
ومن كل ما سبق نستنتج أن التشابه في تركيبة سيتوكروم سي بين الكائنات الحية إنما هو نتيجة انحدارها من أسلاف مشتركة بينها، وكلما كانت الكائنات أبعد في الشجرة التطورية عن بعضها البعض، كلما زادت الاختلافات في تركيبة بروتين سيتوكروم سي، وكل هذا يجعل من سيتوكروم سي أحد أقوى أدلة التطور من الجينوم.
تشفير DNA زائد عن الحاجة
كما هو الحال مع سيتوكروم سي، هناك جينات أخرى واسعة الانتشار. لا تحتاج الكائنات نفس التسلسل الجيني لإنتاج نفس البروتين، ولن نترك المثال السابق، فسنتحدث أيضًا عن الجين المسئول عن إنتاج بروتين سيتوكروم سي.
على الأقل، هناك ثلاث «كودونات-Codons» مختلفة يمكنها إعطاء الأمر للريبوسوم لاختيار نفس الحمض الأميني لوضعه في تسلسل البروتين. (الكودون هو الثلاث حروف من الDNA التي يقرأها الريبوسوم لاختيار الحمض الأميني). وهذا يعني أنه في حالة سيتوكروم سي، هناك حوالي 3¹⁰⁴ أو ما يزيد عن 10⁴⁶ تسلسل جيني مختلف يمكنهم إنتاج نفس البروتين بنفس تسلسل الأحماض الأمينية للقيام بنفس الوظيفة.
وبالتالي فاستخدام الكائنات المختلفة لنفس التسلسل لا يمكن تفسيره وفقًا لفكرة الخلق الخاص ولا التصميم الذكي لكل نوع بشكل منفرد. إذ يمكن لكل كائن أن يستخدم تسلسلًا جينيًا مختلفًا. والعجيب، والمتوقع وفق نظرية التطور، أننا عند فحص التسلسل الجيني المسئول عن إنتاج سيتوكروم سي لكل من الإنسان والشيمبانزي، نجد اختلافًا في 4 نيوكليوتيدات فقط (حوالي 1.2%)، على الرغم من وجود 10⁴⁶ تسلسل جيني ممكن.
في حالة سيتوكروم سي، سنجد أن التشابه في تسلسل الأحماض الأمينية غير ضروري! وكذلك في الجين المسئول عن تشفيره! مما يجعله دليلًا مزدوجًا من أدلة التطور من الجينوم. [15]
«الفيروسات القهقرية-Endogenous Retroviruses»
لا تستطيع الفيروسات التكاثر بذاتها، فعندما يُصاب الكائن بعدوى فيروسية، يقوم الفيروس بإدخال حمضه النووي إلى الحمض النووي الخاص بالخلية، فتقوم الخلية بصنع المزيد من الفيروسات من نفس النوع.
ولكن إذا أصاب الحمض النووي الفيروسي حيوانًا منويًا أو بويضة فسينتقل الحمض النووي الفيروسي إلى الأبناء. هذا لا يعني بالضرورة موت الإبن، إذ أن بعض الطفرات قد تحدث في الجين الفيروسي لتعطيله.
إذا فجينات الفيروسات القهقرية هي جينات موجودة في الجينوم الخاص بك، أتت إليك عندما أصيب أحد أسلافك بهذه الفيروسات ونقل جيناتها إليك على مدى أجيال. تعمل الفيروسات القهقرية كسجل تاريخي لك ولأسلافك من الكائنات الحية التي أصابتها من قبل.
يجب أن تكون الفيروسات القهقرية اختبارًا قويًا لنظرية التطور، أليس كذلك؟ فإذا كنا نحن والرئيسيات الأخرى انحدرنا من سلف مشترك فيجب أن نعثر على جينات فيروسات قهقرية من فيروسات أصابت أسلافهم قبل انفصال الرئيسيات عن بعضها البعض. كما يجب أن نعثر عليها في نفس المكان في الجينوم، إذ أن احتمالية أن يقوم الفيروس بإدخال الجين في نفس المكان على الجينوم لدى نوعين مختلفين دون افتراض وجود علاقة وراثية بينهما هي احتمالية غاية في الضآلة! تخيل أن تراكم عدد هائل من الاحتماليات شديدة الضآلة في كل الكائنات الحية لتفسير الأمر بالصدفة، يصبح تفسيرك مضحك للغاية بحق!
إن كان كل نوع من الرئيسيات قد خُلق بشكل خاص، فلا يجب أن تتشارك الرئيسيات جينات نفس الفيروسات في نفس الأماكن على الحمض النووي الخاص بها بالطبع. تسامحًا مع المنكرين، قد نعثر على نفس الفيروسات، ولكن ليس في نفس الأماكن أبدًا. إذ أن الفيروس عندما يصيب كائنًا ما، فهناك عدد كبير جدًا من الأماكن على الجينوم التي قد يُدخل حمضه النووي فيها. فقد أظهرت دراسات شاملة أن هناك ما يزيد عن 10 مليون مكان ممكن على الجينوم الخاص بالإنسان!
عند مقارنة الجينوم الخاص بالإنسان وذلك الخاص بالشيمبانزي، فاحتمالية عثورنا على جين فيروسي في نفس المكان بين النوعين، وأن يكون الجين عائدًا إلى نفس الفيروس، هي احتمالية أقل من 1 في 10 مليون.
قام العلماء بالبحث عن فيروس HERV-W لدى 12 نوع من الرئيسيات. ويمتلك الشيمبانزي من فيروس HERV-W عدد 208 نسخة، بينما يمتلك الإنسان 211 نسخة، فماذا وجدوا؟
أظهرت الدراسات أن الإنسان والشيمبانزي يتشاركون 205 نسخة من نفس هذا الفيروس في نفس المكان على جينوم كل منهما! [16] أنت الآن أمام دليل لا يدع مجال للشك أمام أي متخصص.
إذا فقد ورث كل من الإنسان والشيمبانزي 205 نسخة من هذا الفيروس من آخر سلف مشترك لهما قبل انفصالهما. بينما ال 6 نسخ الحصرية لدى الإنسان وال 3 نسخ لدى الشيمبانزي، على الأرجح قد ظهرت بعد انفصال المسارات التطورية لكل منهما.
وهذا بحد ذاته كافٍ لتنحية فكرة الخلق الخاص أو التصميم الذكي. إذ أن احتمالية إصابة كلا النوعين بنفس الفيروس في نفس الأماكن على الجينوم بهذا العدد من النسخ بشكل منفصل هو احتمال ضئيل يبلغ 1 في 10¹⁴¹⁸ × 5.88. [17] وهذا يجعل من الفيروسات القهقرية دليلًا بالغ القوة من أدلة التطور من الجينوم.
جين ACYL3
نختتم كلامنا عن أدلة التطور من الجينوم بالحديث عن جين ACYL3 من باب التعديد وليس الإثبات، فما سبق أكثر من كاف. يعمل جين ACYL3 في كل الثدييات على كوكب الأرض (بما في ذلك الغوريلا والأورانجوتان) باستثناء نوعين فقط، وهما الإنسان والشيمبانزي. مما يعني تعطله في آخر سلف مشترك بين الإنسان والشيمبانزي، وهو ما يؤكده كون هذا الجين مُعطل لدى النوعين لنفس السبب بالضبط وهو حدوث طفرة أبدلت الجوانين بالأدينين. فبدلًا من أن يكون التسلسل هو TGG، والذي عند ترجمته يعطي الأمر للريبوسوم باختيار الحمض الأميني «تريبتوفان-Tryptophan»، جعلته الطفرة TGA، والذي عند ترجمته يعطي الأمر للريبوسوم بالتوقف ففسد الجين لدى هذين النوعين فقط. [18]
ولا يمكن تفسير تعطل الجين لدى نوعين فقط بنفس السبب إلا في ضوء نظرية التطور من سلف مشترك. وهو ما يجعله أحد أقوى وأبسط أدلة التطور من الجينوم.
قمنا في هذه السلسلة بسرد عدد كبير من أدلة التطور، وكلها كانت على سبيل المثال لا الحصر. فهناك عدد كبير من الأدلة التي لا يمكننا حصرها في سلسلة مقالات، بل أن هناك أدلة تظهر مع المزيد من الأبحاث يوميًا. فبعض الأدلة الواردة في هذا المقال مأخوذة من أبحاث لم يمر على نشرها 4 سنوات حتى، أي أن الأدلة في تزايد مع استمرار الأبحاث العلمية. فمجال التطور من أكثر المجالات التي تُنشر فيها الأبحاث العلمية. وبعد أن انتهينا من سرد أدلتنا، ننتقل في الجزء القادم إلى تفنيد دعاوى الخلقيين والرد عليهم بخصوص “التصميم الذكي”.
المصادر
[1] The Tech Genetics
[2] Livescience
[3] The Genetic Code: What Is It Good For? An Analysis of the Effects of
Selection Pressures on Genetic Codes, by Olivia P.Judson, Daniel Haydon
[4] Human Origins
[5] PNAS
[6] Springer
[7] Oxford
[8] Inactivation dates of the human and guinea pig vitamin C genes, by Marc Y. Lachapelle, Guy Drouin
[9] Bio med central
[10] Royal Society
[11] Bio med central
[12] PLOS
[13] Pubmed
[14] NCBI
[15] Talk Origins
[16] Pubmed
[17] Google.Docs
[18] PLOS

سعدنا بزيارتك، جميع مقالات الموقع هي ملك موقع الأكاديمية بوست ولا يحق لأي شخص أو جهة استخدامها دون الإشارة إليها كمصدر. تعمل إدارة الموقع على إدارة عملية كتابة المحتوى العلمي دون تدخل مباشر في أسلوب الكاتب، مما يحمل الكاتب المسؤولية عن مدى دقة وسلامة ما يكتب.



التعليقات :